
1. Contexte
La punaise terne Lygus lineolaris (figure 1) est un problème majeur dans les régions productrices de fraises de l'Ontario. Jusqu'à maintenant, le seul moyen de lutte utilisé a été l'application intensive d'insecticides. Les punaises ternes adultes et les larves se nourrissent de toutes les parties de la plante en suçant la sève, en détruisant les embryons des graines et en empêchant la croissance des tissus du fruit sous la graine. Les fruits ainsi déformés sont impropres à la commercialisation (figure 2). En Ontario, la punaise est bivoltine et produit une troisième génération partielle par année dans le sud de l'Ontario. Elle nuit à la production des fraises tout au long de la période de végétation. Les larves de première génération causent des dommages économiques graves aux cultivars produits en juin, et celles de la deuxième génération constituent le principal facteur limitatif des cultivars produits plus tard, qui sont insensibles à la photopériode.
Figure 1. Punaise terne adulte
Agriculture et Agroalimentaire Canada (AAC)

Le risque élevé de développement d'une résistance aux insecticides chez les populations de punaises, la crainte d'effets délétères des insecticides sur les insectes utiles, les pollinisateurs et la santé humaine, ainsi que la concurrence croissante des fraises importées des États-Unis, du Mexique, de la Chine et de l'Europe de l'Est ont motivé la recherche de nouvelles stratégies de lutte contre ces parasites fondées sur la l'absence ou l'utilisation limitée de pesticides classiques.
Figure 2. Dommages causés par la prédation de la punaise terne : fraises difformes.
T. Haye, Centre for Agricultural Bioscience International (CABI)

2. Concept de la lutte intégrée contre les punaises ternes dans les fraises
En 2003, Agriculture et Agroalimentaire Canada (AAC) et l'Agence de réglementation de la lutte antiparasitaire de Santé Canada ont établi le Programme de réduction des risques liés aux pesticides, en collaboration avec le secteur et les provinces, dans le but de réduire les risques liés aux pesticides utilisés en agriculture pour l'environnement et les consommateurs. Dans le cadre de ce Programme, CABI Europe-Suisse, en collaboration avec des partenaires canadiens du ministère de l'Agriculture, de l'Alimentation et des Affaires rurales de l'Ontario, d'AAC et de l'Université de Guelph, a élaboré en 2006 une stratégie de lutte intégrée contre la punaise terne dans les fraiseraies de l'Ontario. La stratégie de lutte intégrée comprend des cultures appâts, des méthodes classiques de lutte biologique et une approche à risque réduit utilisant des moyens de lutte chimique :
- Lorsqu'elles sont plantées le long ou à l'intérieur des champs de fraises, les cultures-appâts telles que la luzerne peuvent limiter les dommages causés par les punaises ternes en leur offrant une source de nourriture qu'elles préfèreront aux fraises. Les punaises adultes provenant des zones avoisinantes se concentrent dans la culture-appât, et le nombre d'adultes qui s'installent dans les fraisiers est réduit. Il a été établi qu'en Californie la luzerne est la culture appât la plus efficace contre les punaises ternes.
- Dans les fraiseraies classiques, on applique des insecticides sur les cultures-appâts après la migration des adultes, ce qui réduit le nombre de punaises ternes femelles avant la ponte. On réduit ainsi le nombre de larves, qui causent le plus de dommages, et l'on évite l'application d'insecticides sur les champs de fraises. Dans les exploitations biologiques, où l'utilisation d'insecticides est interdite, les bandes de luzerne doivent être partiellement coupées et retirées dès que les punaises ternes adultes ont pondu la plupart de leurs oeufs.
- Lorsque l'on note la présence de petites larves de punaises ternes, un agent de lutte biologique classique, le parasitoïde larvaire européen Peristenus digoneutis (figure 3), est lâché pour réduire davantage la population de punaises ternes dans les zones productrices de fraises.
Figure 3. Un Peristenus digoneutis attaque une larve de punaise terne.
T. Haye, CABI

3. Renforcement de la capacité
Afin d'échanger de l'information sur l'organisation et la gestion des fraiseraies en Ontario, la biologie de la punaise terne et les stratégies de lutte de remplacement, on a organisé en 2006 des ateliers participatifs pour les agriculteurs dans des lieux centraux du sud-ouest et de l'est de l'Ontario. Lors de chaque atelier, on a présenté un exposé sommaire décrivant le cycle de vie de la punaise terne, sa migration dans les fraisiers et la façon dont les parasitoïdes interagissent avec la punaise terne et les habitats avoisinants. Au cours des discussions qui ont suivi, des agriculteurs se sont portés volontaires pour l'étude. Comme il y a aussi plusieurs producteurs de fraises biologiques en Ontario, on a décidé que ces derniers seraient les plus ouverts aux nouvelles méthodes de lutte antiparasitaire, et on les a inclus dans l'étude.
En présentant aux producteurs de l'information sur la biologie de la punaise terne et sur la façon dont le parasitoïde P. digoneutis contribue à la mortalité de ce parasite, on a renforcé leur capacité à décider de la meilleure façon de protéger leurs cultures par la lutte biologique. Les producteurs ont acquis des connaissances sur le problème et sur la façon dont une stratégie de lutte biologique peut aider à le résoudre. La discussion sur les contraintes de chaque producteur a permis d'élaborer une stratégie de lutte propre à chaque exploitation. À titre d'exemple, le lieu, le type et l'étendue de la culture-appât sont fonction du terrain disponible.
Après chaque saison de culture, un modèle de rapport du producteur participant a été conçu pour fournir aux producteurs un résumé personnalisé des résultats obtenus dans leur exploitation et les points devant faire l'objet d'un suivi lors des discussions sur le terrain avant la période de végétation suivante. À la fin du projet, on a élaboré un questionnaire visant à évaluer la réaction des producteurs au programme de lutte intégrée.
Figure 4. Lâchés de parasitoïdes dans une exploitation.
K. Makela, AAC

4. Mise en oeuvre de la stratégie de lutte intégrée en Ontario
La mise en œuvre sur le terrain de la stratégie de lutte intégrée en Ontario a commencé en 2007 et s'est poursuivie en 2008. Avec les agriculteurs participants, on a procédé en 2007 et en 2008 à des lâchés périodiques de parasitoïdes larvaires européens P. digoneutis dans les champs de deux sites du sud-ouest de l'Ontario [région de Simcoe] et d'un site de l'est de l'Ontario [région de Kingston] (figure 4). En tout, 8 880 femelles fécondées ont été lâchées directement dans les cultures-appâts adjacentes aux champs de fraises. On a ensuite surveillé la présence de la punaise terne et le parasitisme tout au long de la saison de culture dans les champs de fraises, les cultures-appâts et les zones avoisinantes envahies par les mauvaises herbes (figure 5). Dans le même temps, on a évalué les dommages causés sur les fraisiers afin de mesurer l'incidence de la stratégie de lutte intégrée.
Figure 5. Prise d'échantillons de punaises ternes dans les cultures-appâts.
A. Brauner, AAC

5. Résultats de la mise en œuvre de la stratégie de lutte intégrée
Dans l'est de l'Ontario, ni l'application de pesticides, ni la libération massive de l'agent de lutte biologique P. digoneutis en 2007, n'a empêché l'accroissement de la population de punaises ternes dans tous les sites en 2008 (figure 6). Cette augmentation a probablement été causée par une faible mortalité hivernale des punaises ternes adultes en 2007-2008. On remarque qu'en 2008, on a enregistré la présence du parasitoïde dans des sites où il n'a jamais été lâché, ce qui indique qu'il s'est largement établi dans l'est de l'Ontario. Les données de 2008 montrent clairement que l'augmentation de la densité des punaises ternes s'accompagne d'une augmentation du parasitisme du P. digoneutis (figure 6). Cela concorde avec les observations précédentes faites aux États-Unis, qui montrent que le parasitoïde réagit positivement à l'augmentation de la densité des punaises ternes. L'augmentation dans toute la région du parasitisme des punaises ternes n'a pas encore sensiblement réduit la population de punaises ternes dans l'est de l'Ontario, mais on s'attend à un tel résultat à plus long terme. Dans le sud de l'Ontario, en 2008, on n'a pas observé d'évolution uniforme de la population de punaises ternes (figure 7). Tout comme dans l'est de l'Ontario, les niveaux de parasitisme ont suivi la densité des punaises ternes. Dans les fraiseraies biologiques, où la densité des punaises à chuté en 2008, le parasitisme a également diminué. La tendance contraire a été observée dans la fraiseraie classique où le parasitisme a augmenté en même temps que la densité des punaises. La différence entre les dommages causés par la punaise terne en 2007 et en 2008 dans les fraiseraies classiques a tantôt augmenté tantôt diminué sur le site, probablement en raison du recours à des techniques de lutte différentes de celles des années précédentes (figure 8). En revanche, les dommages causés aux fraises sont restés les mêmes durant ces années dans les deux exploitations biologiques participant au programme de lutte intégrée. Globalement, trois exploitants n'ont pas appliqué de pesticides, et une exploitation en a largement réduit son utilisation en 2008 par rapport aux années précédentes.
Figure 6. Nombre total de larves aux stade N4 et N5 pendant la saison de récolte et pourcentage de parasitisme à la date où leur nombre était le plus élevé dans les fraiseraies de l'est de l'Ontario en 2007 et en 2008. * Indique un changement important du nombre de larves (Tukey Test, P < 0.05).
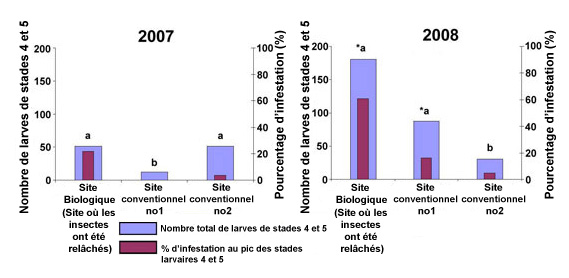
Description - Figure 6
Deux graphiques à barres présentant les années 2007 et 2008.
Chaque graphique indique le nombre de larves de stades 4 et 5 (échelle 0–200) et le pourcentage d'infestation (échelle 0–100 %). Chaque graphique présente trois sites : le site biologique (site de lâcher de parasitoïdes), le site conventionnel no 1 et le site conventionnel no 2. Chaque graphique contient deux variables : le nombre total de larves de stade 4 et 5 et le pic des pourcentages d'infestation des stades larvaires 4 et 5.
En 2007, le pic des pourcentages d'infestation a été en moyenne d'environ 20 % au site biologique, de 0 % au site conventionnel no 1 et de moins de 5 % au site conventionnel no 2. Le nombre total de larves s'est élevé à environ 50 au site biologique, à moins de 10 au site conventionnel no 1 et à environ 50 au site conventionnel no 2.
En 2008, le pourcentage d'infestation des fraiseraies a été plus élevé partout, dépassant 60 % au site biologique et atteignant près de 20 % au site conventionnel no 1 et près de 5 % au site conventionnel no 2. Le nombre total de larves avoisinait 170 au site biologique, 100 au site conventionnel no 1 et 40 au site conventionnel no 2. Le pourcentage d'infestation était plus élevé dans les sites où le nombre de larves de stades 4 et 5 avait augmenté. Pour les deux années, le pourcentage d'infestation du site biologique a été significativement plus élevé que celui des sites conventionnels.
Figure 7. Nombre total de larves aux stade N4 et N5 pendant la saison de récolte et pourcentage de parasitisme à la date où leur nombre était le plus élevé dans les fraiseraies du sud de l'Ontario en 2007 et en 2008. * Indique un changement important du nombre de larves aux stade N4 et N5 entre ces deux années; les nombres de larves aux stade N4 et N5 suivis de la même lettre ne sont pas très différents (test de Tukey, P < 0.05).
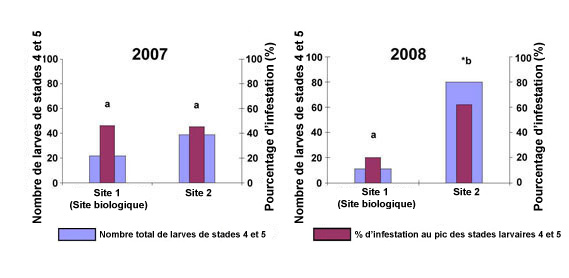
Description - Figure 7
Deux graphiques à barres présentant les années 2007 et 2008.
Chaque graphique indique le nombre de larves de stades 4 et 5 (échelle 0–100) et le pourcentage d'infestation (échelle 0–100 %) au site biologique no 1 et au site conventionnel no 2. Chaque graphique contient deux variables : le nombre total de larves de stade 4 et 5 et le pic des pourcentages d'infestation des stades larvaires 4 et 5.
En 2007, les pourcentages d'infestation du site biologique ont culminé en moyenne à plus de 40 % et ceux du site conventionnel à près de 40 %. Environ 20 larves de stades 4 et 5 ont été dénombrées au site biologique et 40 au site conventionnel.
En 2008, le pourcentage moyen d'infestation du site biologique a été plus bas (20 %) qu'en 2007 et celui du site conventionnel, plus élevé (60 %). Ces données correspondent à la diminution du nombre de larves de punaises ternes au site biologique (environ 10) et à l'augmentation du nombre de larves de stades 4 et 5 au site conventionnel (environ 80).
Figure 8. Variation des dommages causés par la punaise terne aux fraises entre 2007 et 2008 dans les exploitations de l'est de l'Ontario (3 récoltes/site/année). * Indique une variation importante des dommages généraux causés par la punaise terne pendant ces années; globalement, les dommages causés par la punaise terne aux fraises suivis de la même lettre ne présentent pas de grande différence (test de Tukey P < 0.05).
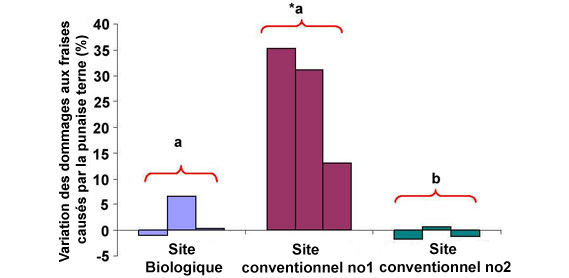
Description - Figure 8
Graphique à une barre représentant le pourcentage de variation des fruits endommagés par les larves de stades 4 et 5 entre 2007 et 2008.
L'axe vertical du graphique montre la variation du pourcentage de fruits endommagés (échelle de −5 % à 40 %). Chaque graphique présente trois sites : le site biologique (site de lâcher de parasitoïdes), le site conventionnel no 1 et le site conventionnel no 2. Chaque site est représenté par trois barres qui correspondent à différentes dates de cueillette. Au site conventionnel no 1, le pourcentage de fruits endommagés en 2008 a été significativement supérieur à celui en 2007 (+35 %, +30 % et +12 %). Il n'y a aucune différence significative du pourcentage de fruits endommagés entre les deux années au site biologique (−1 %, +9 %, presque 0 %) et au site conventionnel no 2 (−4 %, +1 %, 2 %).
6. Conclusions et perspectives d'avenir
La distribution actuelle du P. digoneutis au Canada montre que ce parasitoïde est désormais bien établi dans l'est de l'Ontario et dans certaines régions du sud de l'Ontario. Dans le nord-est des États-Unis, il a fallu presque dix ans avant que l'on remarque une incidence importante du P. digoneutis et que la population de punaises ternes disparaisse. On espère donc qu'une croissance continue de la population de P. digoneutis en Ontario aura des effets similaires à long terme et engendrera une réduction du nombre de parasites dans toute la région. Cela préconise un travail continu visant à favoriser la croissance de la population et à évaluer les effets de la stratégie. En outre, en réalisant des sondages dans d'autres régions de l'Ontario, on pourrait cerner les zones où le parasite n'est pas encore établi, et des lâchés pourraient accélérer la dispersion et l'établissement du P. digoneutis.
Au sujet du Programme de réduction des risques liés aux pesticides d'Agriculture et Agroalimentaire Canada
L'équipe de réduction des risques liés aux pesticides offre aux producteurs canadiens des solutions viables pour réduire les risques liés aux pesticides dans l'industrie agricole et agroalimentaire. L'équipe accomplit cet objectif en finançant des projets de lutte intégrée et en coordonnant des stratégies de réduction des risques liés aux pesticides élaborées en consultation avec les intervenants et les experts en lutte antiparasitaire. D'autres fiches d'information sur la protection durable des cultures sont disponibles. Pour plus d'informations, veuillez visiter le Centre de la lutte antiparasitaire.